
Генотип как целостная система
В начале ХХ века генетик из Дании — Вильгельм Иогансен впервые предложил именовать термином «генотип» наследственные признаки человека. Им же были введены такие термины как: «популяция», «чистые линии», «фенотип», «аллель» и «ген».
Определение понятия
Генотипом именуют все гены, находящиеся внутри конкретного живого существа. Согласно статистическим исследованиям, в человеке находится не менее 35000 генов. Единой функциональной системой генотип стал в результате происходящих эволюционных изменений. Процесс генного взаимодействия считается признаком системности генотипа.
Всем генам присущи следующие свойства:
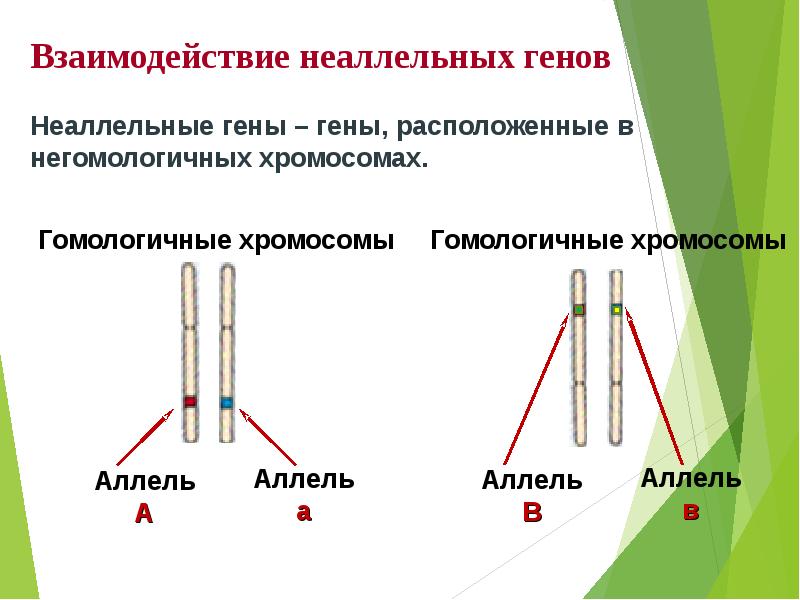
- взаимодействие с другими неаллельными единицами наследственности;
- дискретностьдействий;
- частота встречаемости (пенетрантность);
- степени выражения признаков (экспрессивность);
- передача неизмененных наследственных свойств (стабильность);
- ответственность одного гена за несколько признаков (плейотропность);
- специфичность;
- неустойчивость к мутационным изменениям (лабильность).
Из всего выше перечисленного можно сделать вывод, что генотипом является целостный генетический аппарат, компоненты которого взаимодействуют друг с другом.
Связь генотипа и признаков
Г. Мендель впервые разработал главные закономерности наследственности, доказывая вероятность наследования одного признака независимо от других. Так, была определена делимость наследственности, а в генотипе выделили отдельные единицы, определяющие отдельные признаки.
Позднее, стало понятно, что одному и тому же гену свойственно влияние на несколько разных признаков, находящихся в организме. Взаимодействие генов также влияет на проявление разных признаков в организме. Таким образом, генотип стал считаться целостной системой живого.
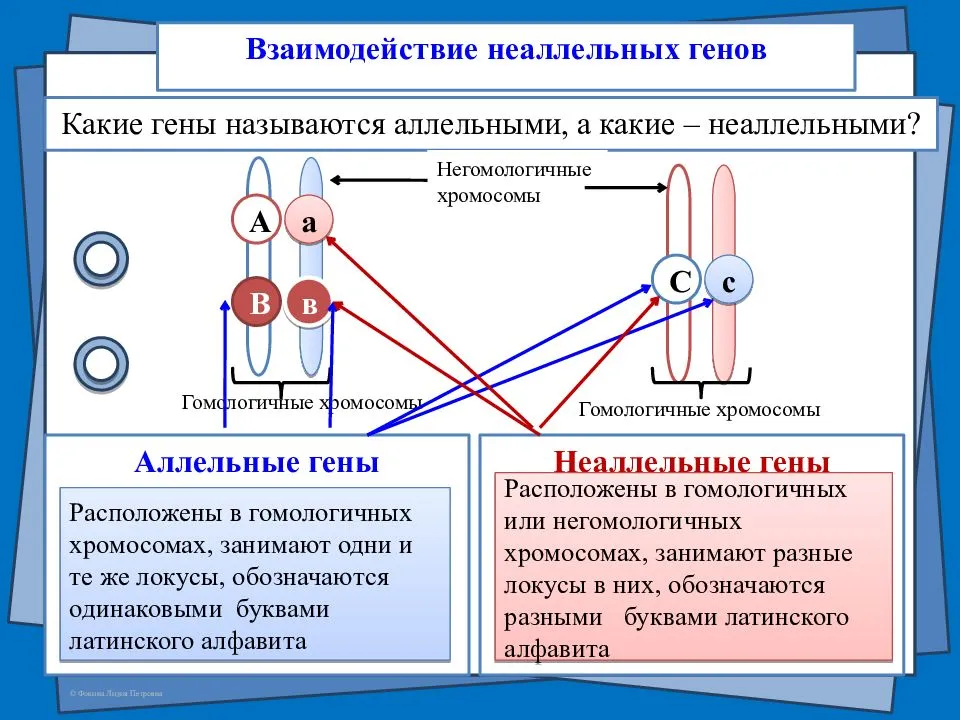
Взаимодействие между аллельными генами
Генотип и фенотип. Взаимодействие аллельных генов.
Фенотип — это весь комплекс внешних и внутренних признаков организма. Например, размер, цвет, поведение, активность ферментов.
Генотип — это система генов организма, взаимодействующих между собой. Взаимодействие генов может менять наследование определенных признаков.
Взаимодействуют гены на разных уровнях:
1. На уровне генома (наследственного материала).
2. На уровне продуктов функциональной активности генов (белков-ферментов, РНК…).
Взаимодействие между аллельными генами.
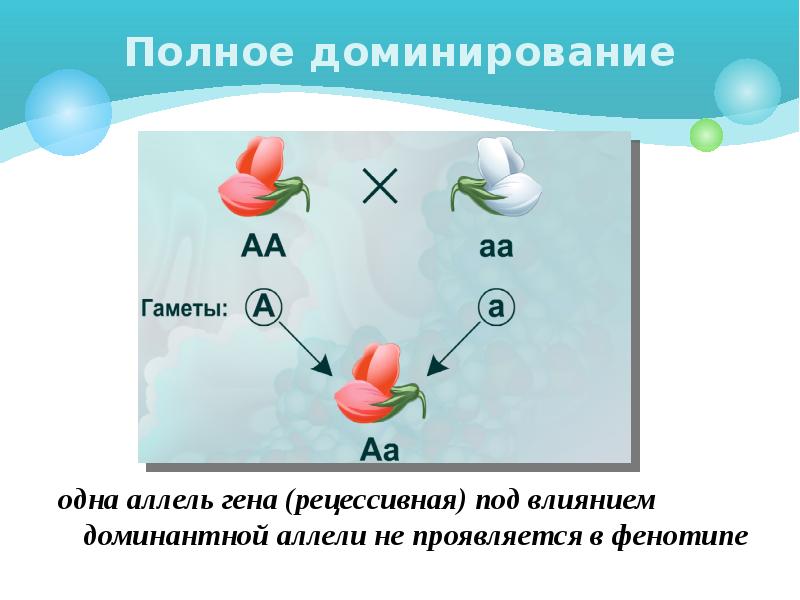
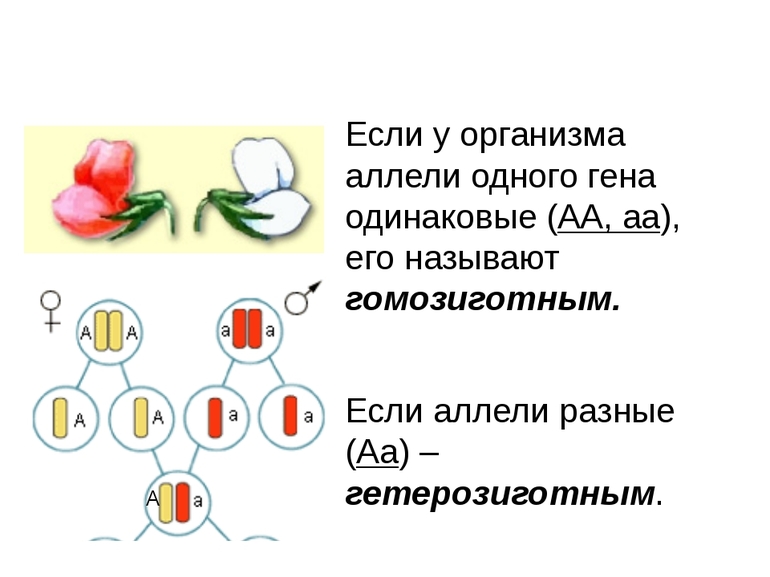
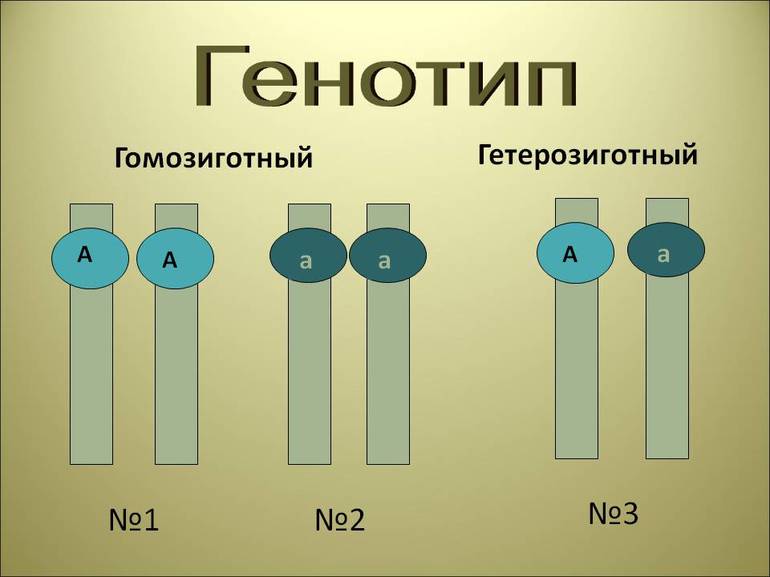
1. Доминантность и рецессивность. Доминантный ген проявляет себя в признак как в гомозиготном, так и в гетерозиготном состоянии.
Рецессивный ген проявляет себя в признак только в гомозиготном состоянии. Например: у человека ген карего цвета глаз доминирует над голубым, у мушки дрозофилы ген, отвечающий за длинные крылья доминирует над геном
коротких крыльев.
3. Кодоминантность — это такая форма взаимодействия, когда оба аллеля одного гена проявляют себя фенотипически независимо друг от друга.
Например: наследование IV (АВ) группы крови у человека, когда аллели IА, IВ одновременно проявляют себя в признак.
За гемоглобин крови у человека отвечает НЬА-аллель. Если происходит мутация и в организме появляется HbS-аллель, то в организме одновременно присутствуют нормальные эритроциты и эритроциты в виде серпа.
Относительное доминирование — это такое взаимодействие, когда проявление гена зависит от условий, т. е. в одних условиях ген ведет себя как доминантный, а в других как рецессивный. Например, ген отвечающий за облысение в мужском организме на фоне мужских половых гормонов ведет себя как доминантный, а в женском организме на фоне женских половых гормонов — как рецессивный.
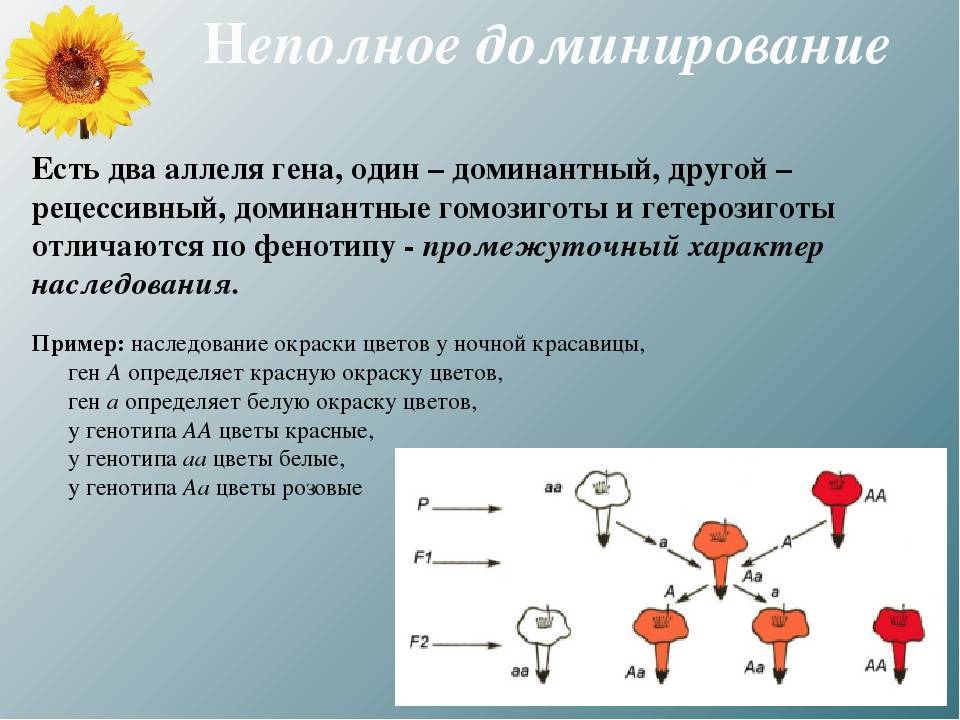
4. Неполное доминирование — такая форма взаимодействия, когда в гетерозиготном состоянии (Аа) проявляется среднее значение признака между доминантным и рецессивным.
Например, наследование окраски цвет ков львиного зева:
А-ген, отвечающий за красную окраскуцветков
а — за белую окраску
Аа — генотип растения с розовыми цветками.
Р
Г А а
F1 Аа все растения имеют розовые цветки
Например: у человека заболевание фенилкетонурия связано с рецессивным мутантным геном.
В гомозиготном состоянии (аа) активность фермента, который превращает фенилаланин в тирозин равна 0%. В гомозиготном состоянии (АА) — активность фермента 100%. А в гетерозиготном состоянии (Аа) активность фермента 50%.


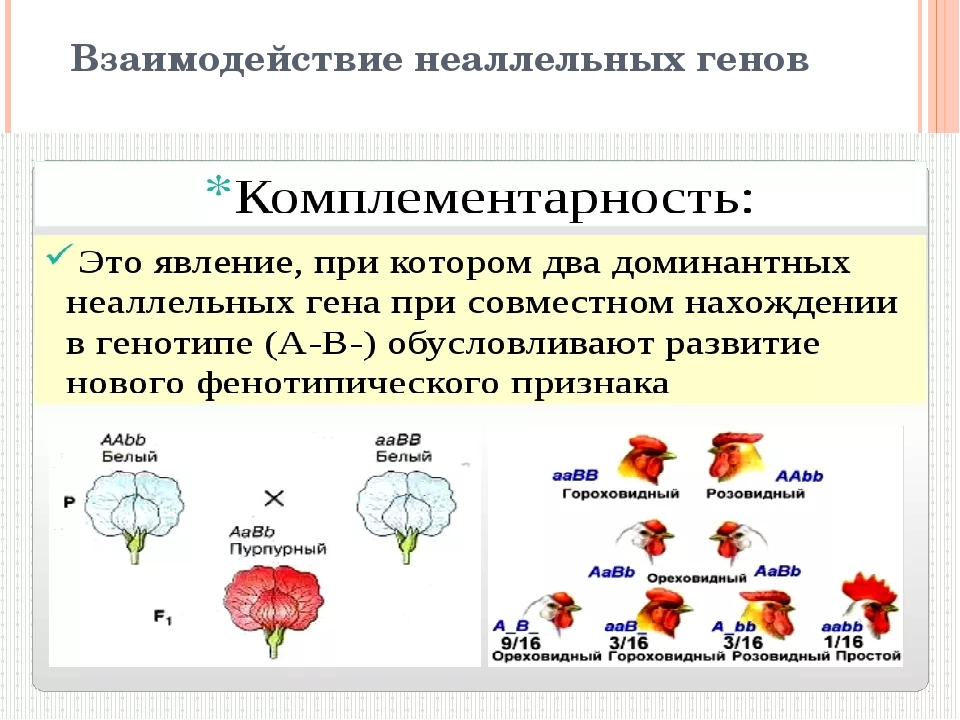


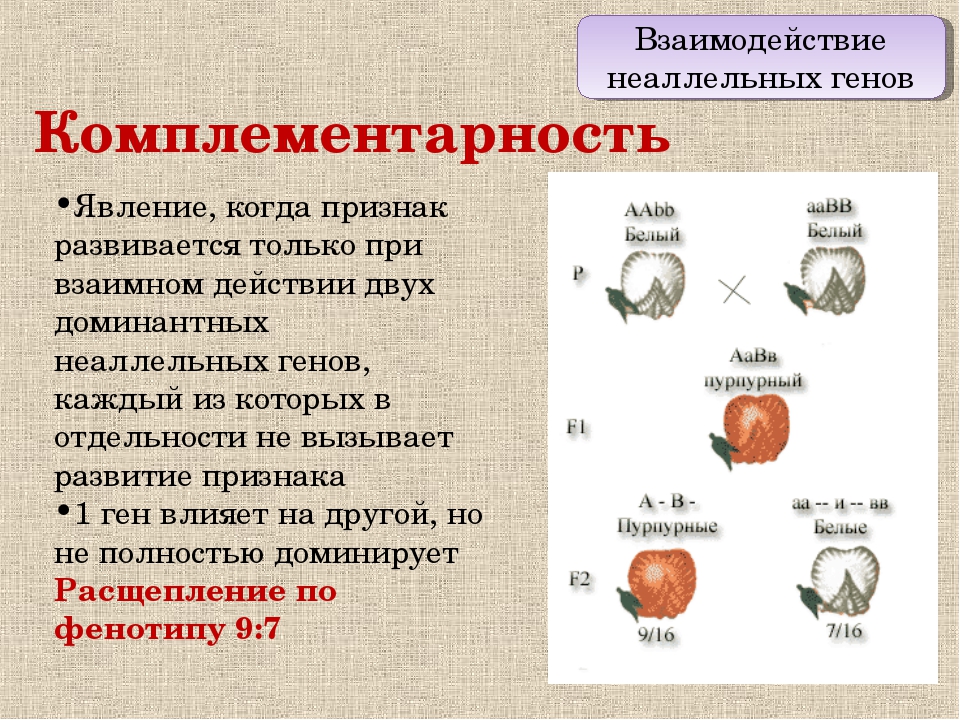
Множественные аллели. Наследование групп крови АВО.
Гены могут мутировать в несколько состояний:
А →А1 → А2→А3…
Мутации одного гена во много состояний называется серией множественных аллелей.
В серии множественных аллелей 3 и более генов.
Генетика человека
Генетикой человека именуют одно из направлений биологии, которое занимается изучением закономерностей наследования, эволюции и происходящих в генотипе изменений. Основной задачей данной науки является определение генотипа и условий его формирования.
Общие представления
Генетика человека основывается на общих закономерностях наследования признаков, затрагивая разные биологические области:
- физиология;
- биохимия;
- морфология.
Ученым удалось выделить и детально изучить более 20 типов генетических сцеплений, а также они смогли составить хромосомные карты, фиксируя на них разные гены. Многочисленные исследования малочисленных групп людей, где высока частота заключения браков, доказали скорое родство всех людей такого социума. Это весьма опасно, так как приводит к повышению риска развития рецессивных мутаций. Обычно такие беременности заканчиваются летальным исходом.
Будущие перспективы
Основными направлениями деятельности при изучении человеческой генетики являются механизмы выявления наследования признаков у разных рас. Выяснено, что от человека к человеку происходит существенное изменение набора имеющихся признаков. При этом получают объяснения следующие виды наследования:
- сцепленное с полом;
- доминантное;
- кодоминантное;
- рецессивное;
- аутосомное.
С целью достижения максимальной точности производимых исследований, необходима разработка и применение специфических методов, которые сегодня широко внедряются в практику. Применение аналитических подходов в человеческой генетике, основанных на анализе имеющейся информации, помогает расширить изученный каталог человеческих признаков, передающихся из поколения в поколение. Нередко, генетику человека именуют «антропогенетикой».
Смотри также:
- Закономерности наследования, установленные Г. Менделем, их цитологические основы (моно- и дигибридное скрещивание)
- Законы Т. Моргана: сцепленное наследование признаков, нарушение сцепления генов
- Закономерности изменчивости. Ненаследственная (модификационная) изменчивость. Норма реакции
Наследование признаков, сцепленных с полом
Наследованием, сцепленным с полом, именуют порядок наследования генов, которые находятся внутри половых хромосом. Сцепленное с X-хромосомой наследование представляет собой процесс наследования генов гетерогаметного мужского пола, имеющего Y-хромосому(XY) и гомогаметной женской особи (XX). Данный тип передачи генов работает у:
- пресмыкающихся;
- многих насекомых;
- млекопитающих (кроме однопроходных).
Если наследование признаков осуществляется посредством Y-хромосомы, имеет место голандрический тип передачи признаков. При голандрическом наследовании происходит передача признаков, гены которых локализованы только в Y-хромосоме. Так передаются к сыновьям от отцов фенотипические проявления. Данными признаками служат:
- ген дифференцировки семенников;
- наличие межпальцевых перепонок;
- развитие волос по краю ушной раковины (гипертрихоз).
Гипертрихоз — это болезнь, вызывающая у человека чрезмерный рост волос. Волосы являются производными верхнего слоя кожи человека и млекопитающих. Их корни располагаются в толще кожи. До 8 месяцев утробного развития человека его тело покрыто плодными и первичными волосами, но к моменту рождения волосы остаются только на бровях, ресницах и голове.
Гомозиготными признаками именуют проявления, гены которых локализованы только в одной (X либо Y) половой хромосоме. Таким признакам не свойственно наличие аллельных генов в других половых хромосомах. Примером человеческого недуга, сцепленного с полом, служит дальтонизм, ген которого доминирует над геном, отвечающим за нормальное зрение.
Основные типы отношений между организмами
Симбиотические отношения

Симбиоз (+ / +) описывает тесное и постоянное (долгосрочное) взаимодействие между двумя видами.
Симбиотические отношения подразделяются на облигатные (нужны для выживания) или факультативные (выгодные, но не строго необходимые).
Симбиотические отношения могут быть полезны как для одного, так и для обоих организмов в партнерстве:
- Мутуализм (+ / +) – оба вида извлекают выгоду от взаимодействия (анемон защищает рыбу-клоуна, рыба-клоун обеспечивает анемон питательными фекалиями);
- Комменсализм (0 / +) – один вид получает выгоду, а другой остается невредим (ракообразные прикрепляются к китам, чтобы добраться до мест богатых планктоном);
- Паразитизм (– / +) – один из видов получает выгоду в ущерб другому (например, клещи и блохи питаются кровью своего хозяина).
Хищник-Жертва
Хищничество (– / +) – это биологическое взаимодействие, при котором один организм (хищник) охотится и питается другим организмом (добычей).
Поскольку хищник полагается на добычу как на источник пищи, численность их популяций неразрывно связана между собой:
- Если численность популяции добычи сокращается (например, из-за чрезмерной охоты), численность хищников будет сокращаться по мере усиления внутривидовой конкуренции;
- Если численность популяция добычи растет, численность хищников будет увеличиваться в результате изобилия источника пищи.
Нейтрализм

Нейтрализм (0 / 0) описывает взаимосвязь между двумя видами , которые взаимодействуют, но никак не влияют друг на друга. Примеры истинного нейтрализму практически невозможно доказать. На практике термин используется для описания ситуаций, где взаимодействия пренебрежимо малы или незначительны.
Аменсализм

Аменсализм (– / 0) – межвидовые отношения, при которых одному организму наноситься вред, а другой остается невредимым. Хорошим примером аменсализма является крупный рогатый скот, который во время передвижения затаптывает копытами траву.
Конкуренция

Конкуренция (– / –) описывает взаимодействие между двумя организмами, при котором приспособленность одного снижается из-за присутствия другого.
Конкуренция может быть внутривидовой (между представителями одного вида) или межвидовой (между представителями разных видов).
Нехватка ресурсов (например, еды, воды или территории) обычно вызывают один из двух типов конкуренции:
- Конкурентное исключение – один вид использует ресурсы более эффективно, приводя к сокращению популяции других видов;
- Разделение ресурсов – оба вида разделяют ресурсы в общей среде обитания, при этом не нанося существенного вреда друг другу.
7.Множественный аллелизм. Генетика групп крови.
Множественный
аллелизм — различные состояния (три и
более) одного и того же локуса хромосом,
возникшие в результате мутаций.
Присутствие
в генофонде вида одновременно различных
аллелей гена называют множественным
аллелизмом. Примером этому служат разные
варианты окраски глаз у плодовой мухи:
белая, вишневая,
красная, абрикосовая,
эозиновая,— обусловленные различными
аллелями соответствующего гена. У
человека, как и у других представителей
органического мира, множественный
аллелизм свойствен многим генам. Так,
три аллеля гена I определяют групповую
принадлежность крови по системе АВ0
(IA, IB, I0). Два аллеля имеет ген, обусловливающий
резус-принадлежность. Более ста аллелей
насчитывают гены α- и β-полипептидов
гемоглобина.
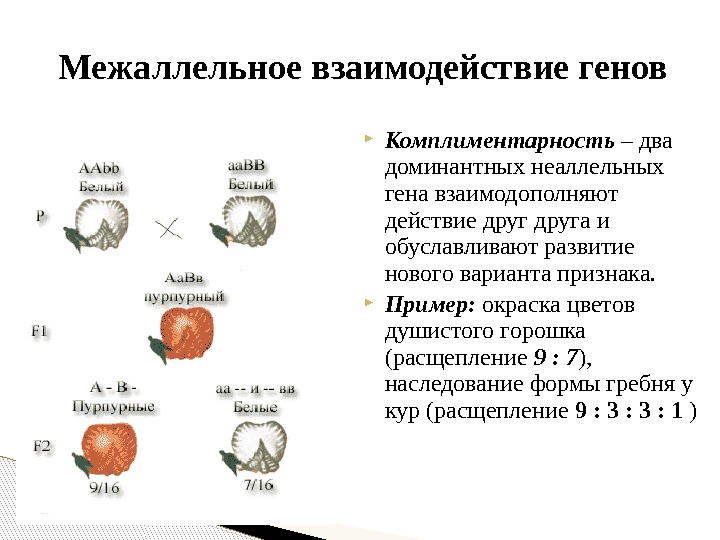
Система
групп крови АВО
Система
групп крови ABO — это основная система
групп крови, которая используется при
переливании крови у людей. Ассоциированные
анти-А и анти-В-антитела (иммуноглобулины),
обычно относятся к типу IgM, которые, как
правило, образуются в первые годы жизни
в процессе сенситизации к веществам,
которые находятся вокруг, в основном
таких, как продукты питания, бактерии
и вирусы.
Система групп крови ABO также присутствует
у некоторых животных, например, у обезьян
(шимпанзе, бонобо и горилл).
Наследование
групп крови системы АВО. В системе АВО
синтез агглютиногенов и агглютининов
определяется аллелями гена I:I0, IA, IB. Ген
I контролирует и образование антигенов,
и образование антител.
При этом наблюдается полное доминирование
аллелей IA и Ю над аллелем I0, но совместное
доминирование (кодоминирование) аллелейIА
и IB. Из-за кодоминирования наследование
групп крови системы АВО происходит
сложным образом. Например, если мать
гетерозиготна по II группе, крови (генотип
IAI0), а отец гетерозиготен по Ш группе
крови (генотип IBI0), то в их потомстве с
равной вероятностью может родиться
ребенок с любой группой крови. Если у
матери I группа крови (генотип I0I0), а у
отца IV группа крови (генотип IАIB), то в
их потомстве с равной вероятностью
может родиться ребенок или со П (генотип
IAI0), или с III (генотип IBI0) группой крови
(но не с I, и не с IV).
Цитоплазматическая наследственность
Цитоплазматическая наследственность — наследственность, связанная с действием генов, находящихся в органоидах цитоплазмы, содержащих ДНК (митохондриях и пластидах).
■ Такие гены способны к автономной репликации и равномерному распределению между дочерними клетками.
■ В передаче признаков цитоплазматическая наследственность имеет второстепенное значение.
■ Цитоплазматическое наследование осуществляется только через материнский организм (в мужских половых клетках цитоплазмы мало и в ней митохондрии и пластиды отсутствуют). Примеры: с мутациями генов, локализованных в митохондриях, связано наследование нарушений в действии дыхательных ферментов у дрожжей; с мутациями генов, локализованных в пластидах, связано, в частности, наследование пестролистности у ряда растений (ночной красавицы, львиного зева и др.).
генетика
Причины отклонений от менделевских расщеплений
Менделевские расщепления в принципе могут быть реализованы только для видов живых организмов с нормальным половым процессом, диплоидностью обоих полов и при условии отсутствия нарушений в мейозе. Но и при выполнении указанных условий случаи отклонения от менделевских расщеплений достаточно многочисленны. При этом каких-либо исключений нет, а условия менделирования не нарушаются — просто на них накладываются другие эффекты.

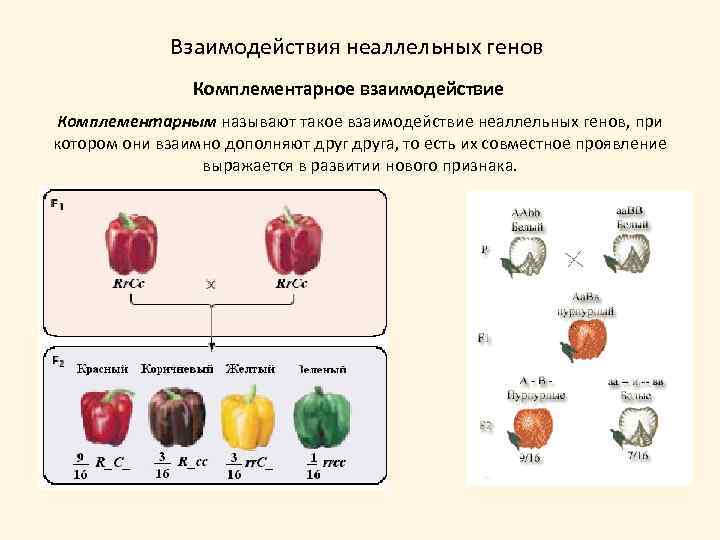
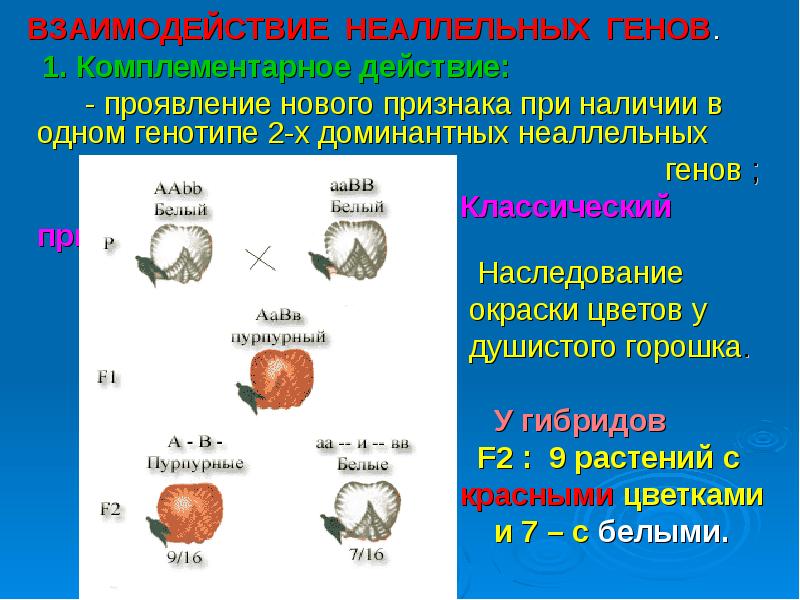
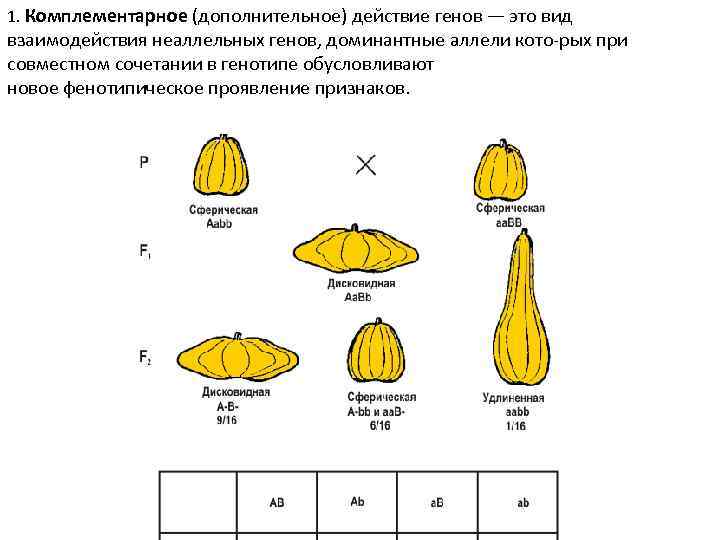
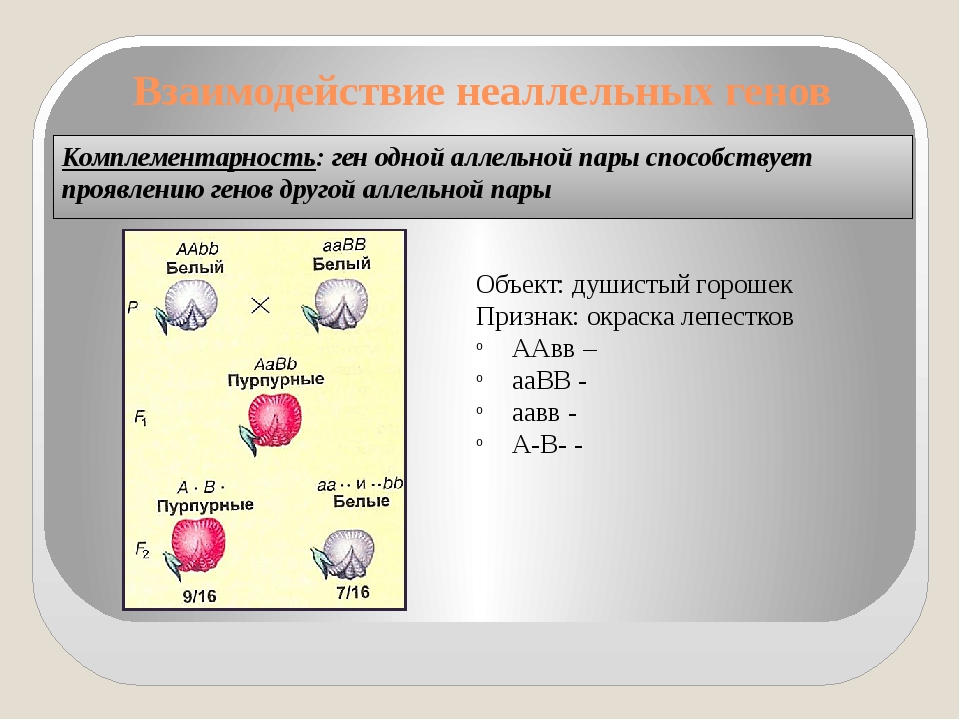
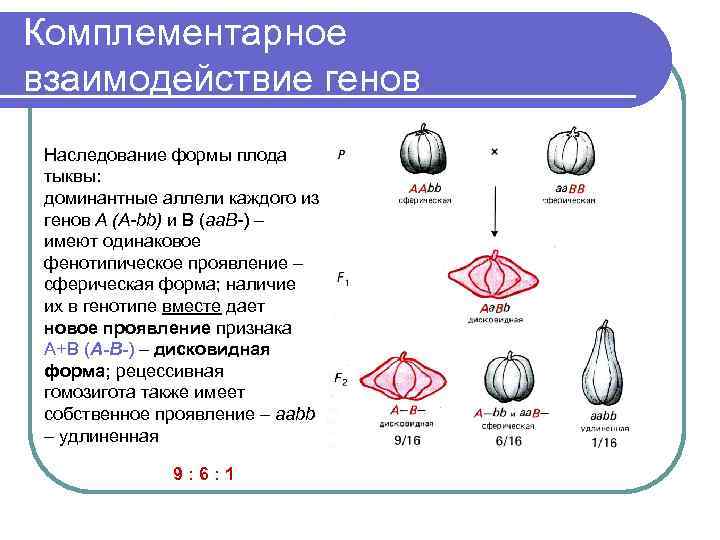
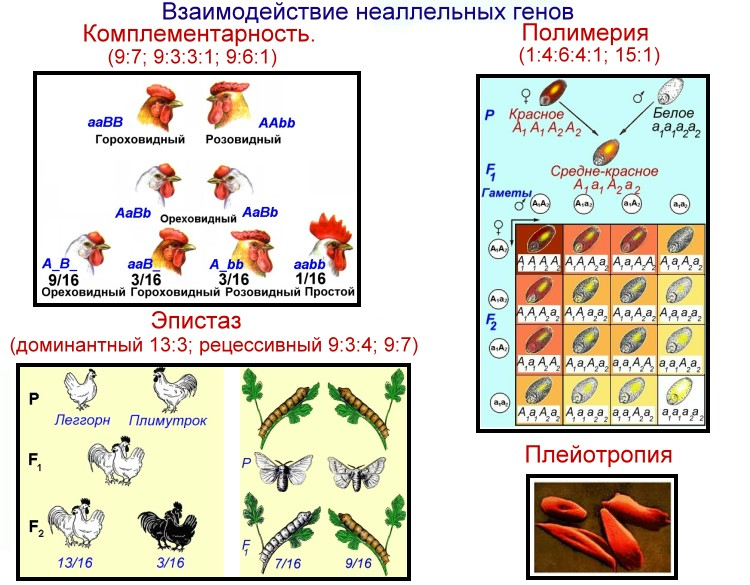
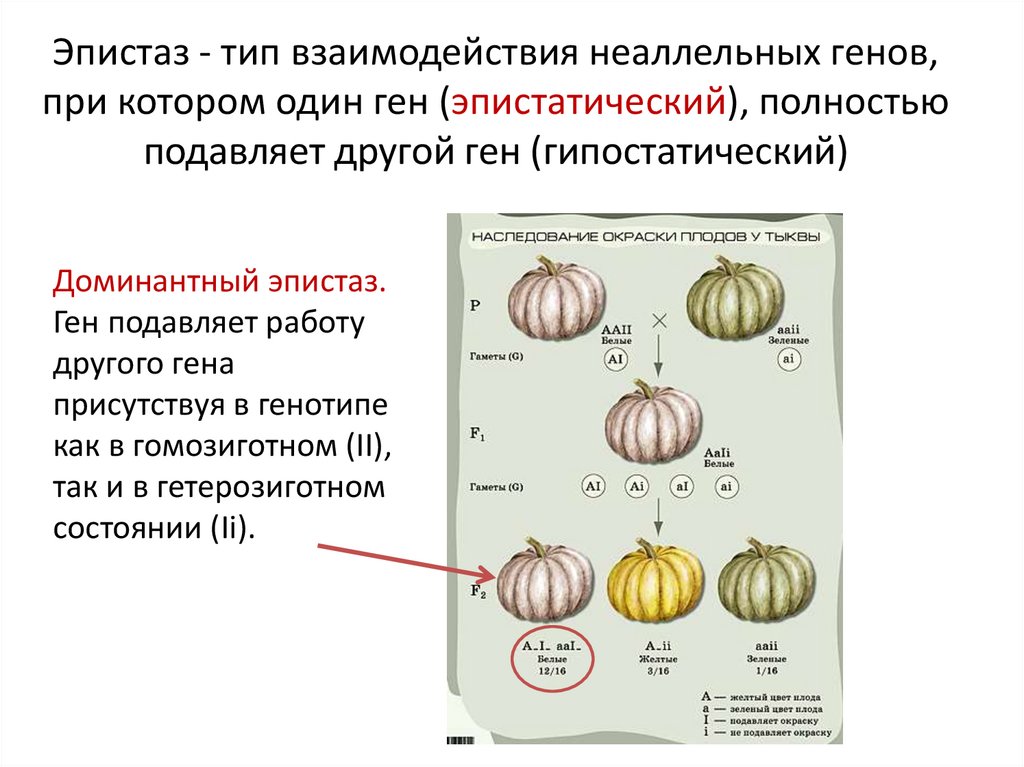
Три таких эффекта — взаимодействие неаллельных генов (комплементация, кумулятивная и некумулятивная полимерия), сцепление групп генов в одной хромосоме и расположение гена в половых хромосомах. Среди других причин выделяются такие:
- Летальность некоторых комбинаций аллелей. Например, при скрещивании жёлтых (гетерозиготных) мышей, расщепление на жёлтые и чёрные (рецессивные гомозиготы) всегда составляет 2:1. Подобный феномен наблюдается также при скрещивании платиновых лисиц. Если проверить беременных самок, то оказывается, что четверть эмбрионов погибает ещё до рождения: погибают гомозиготы по доминантному аллелю, поэтому и наблюдается отклонение от ожидаемого расщепления 3:1 (отсутствует один из четырёх генотипических классов). Таким образом, в этой задаче доминантный аллель является на самом деле доминантным только по окрасу шерсти. Что касается жизнеспособности, этот самый аллель является рецессивным. Ситуация, когда один ген влияет на несколько признаков одновременно (окрас шерсти и жизнеспособность), является одним из примеров плейотропии.
- Существуют аллели, которые фенотипически проявляются только у определённой группы организмов, содержащих их в генотипе. Для характеристики такого вариабельного проявления генов используют понятие пенетрантности и экспрессивности. Согласно определению, мерой пенетрантности является группа особей, характеризующихся определённым фенотипом, среди всех особей с одинаковым генотипом. Некоторые наследственные болезни человека, например, подагра, характеризуются неполной пенетрантностью, то есть не все особи, которые несут мутацию, болеют. Экспрессивность отображает силу проявления мутантного гена в фенотипе. Так, дрозофилы, гомозиготные по мутантному аллелю eyeless, могут иметь разное количество фасеток глаз — от нормального количества до полного их отсутствия. Способность генотипа проявляться по-разному в зависимости от внешних условий отражает норму реакции — определённый диапазон проявления (изменчивость), в границах которого признаки могут модифицироваться в ответ на вариации условий развития организма.
- Некоторые признаки присущи особям в течение не всей жизни, а возникают только в определённый период: характер расщепления может иметь взаимосвязь с возрастом. Например, у человека имеются наследственные синдромы, которые проявляются только в преклонном возрасте (синдром Альцгеймера, болезнь Хантингтона).
- К отклонениям от ожидаемых менделевских расщеплений приводит также явление импринтинга. Основой его является так называемая эпигенетическая наследственность — может наследоваться от родителей не просто ДНК, а хроматина, который несёт определённые химические маркеры. От них зависит структурное состояние участка и, соответственно, активность некоторых генов. Вследствие такого эффекта проявление определённого гена может быть обусловлено тем, от кого из родителей хромосома была получена.
- В природе существует много видов полового диморфизма, и пол играет большую роль в проявлении ряда признаков.
Новая форма наследования
Иногда в результате скрещивания потомки наследовали промежуточные признаки, которые не давал в гомозиготной форме родительский ген. Неполного доминирования не было в понятийном аппарате генетики до начала XX века, когда были переоткрыты законы Менделя. Тогда же многие естествоиспытатели проводили генетические эксперименты с растительными и животными объектами (томатами, бобовыми, хомяками, мышами, дрозофилами).
После цитологического подтверждения в 1902 г. Уолтером Сеттоном Менделевских закономерностей принципы передачи и взаимодействия признаков стали объяснять с позиции поведения хромосом в клетке.

В том же 1902 году Чермак Корренс описал случай, когда после скрещивания растений с белыми и красными венчиками у потомков были цветки розовой окраски – неполное доминирование. Это проявление у гибридов (генотип Аа) признака, который является промежуточным по отношению к гомозиготному доминантному (АА) и рецессивному (аа) фенотипам. Подобный эффект описан для многих видов цветковых растений: львиного зева, гиацинта, ночной красавицы, ягод земляники.
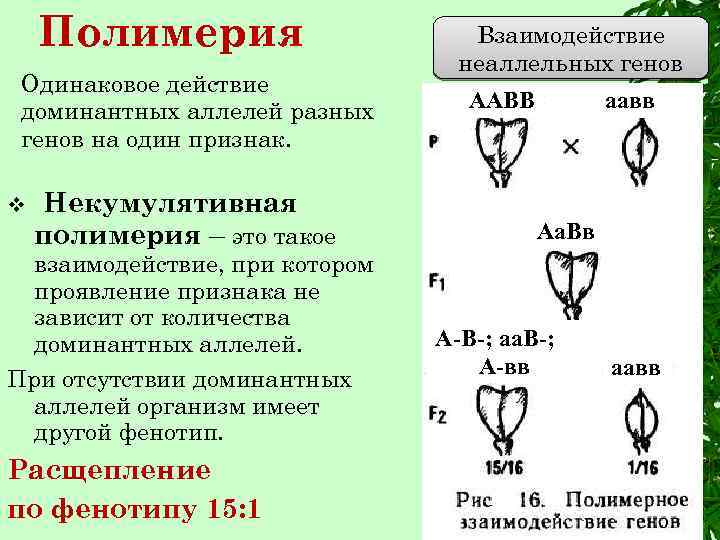
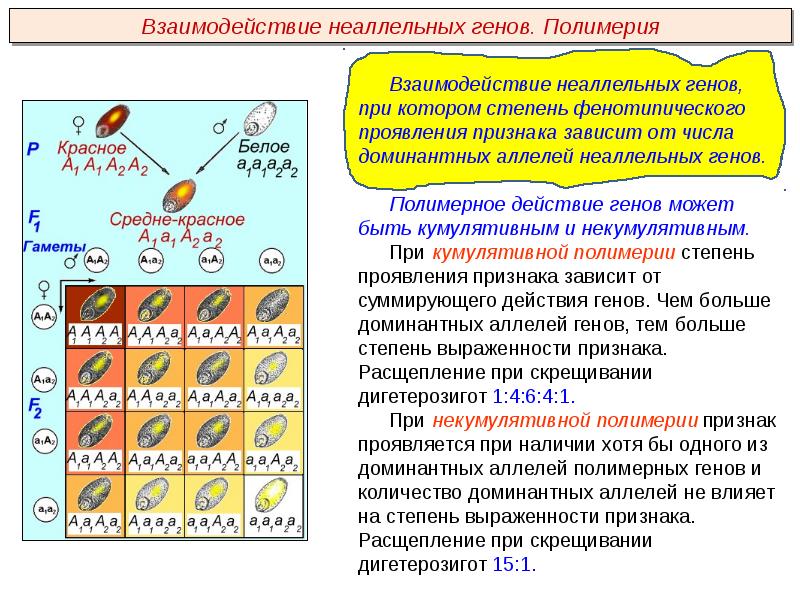
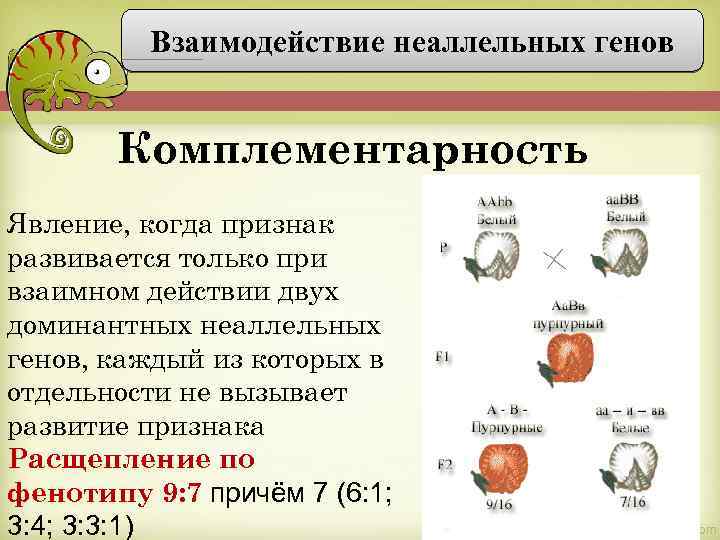
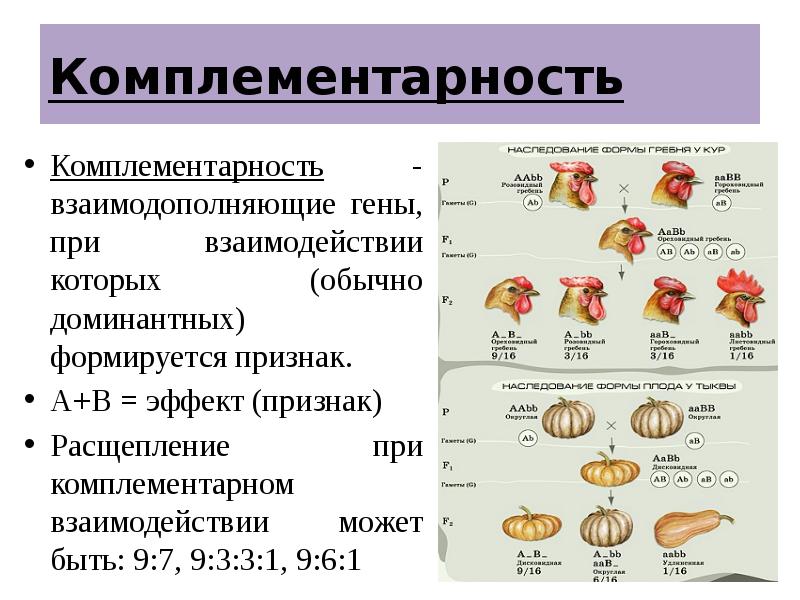
Полимерия
Это вид взаимодействия двух и более пар неаллельных генов, доминантные аллели которых однозначно влияют на развитие одного и того же признака. Полимерное действие генов может быть кумулятивным и некумулятивным. При кумулятивной полимерии интенсивность значения признака зависит от суммирующего действия генов: чем больше доминантных аллелей, тем больше степень выраженности признака. При некумулятивной полимерии количество доминантных аллелей на степень выраженности признака не влияет, и признак проявляется при наличии хотя бы одного из доминантных аллелей. Полимерные гены обозначаются одной буквой, аллели одного локуса имеют одинаковый цифровой индекс, например А1а1А2а2А3а3.
Кумулятивная полимерия имеет место при наследовании окраски зерновок пшеницы, чешуек семян овса, роста и цвета кожи человека и т.д.
| P | A1A1A2A2черные чешуйки | × | a1a1a2a2белые чешуйки |
| Типы гамет | A1A2 | a1a2 | |
| F1 | A1a1A2a2серые чешуйки, 100% | ||
| P | A1a1A2a2серые чешуйки | × | A1a1A2a2серые чешуйки |
| Типы гамет | A1A2 A1a2 a1A2 a1a2 | A1A2 A1a2 a1A2 a1a2 |
| A1A2 | A1a2 | a1A2 | a1a2 | ||
| A1A2 | A1A1A2A2черные | A1A1A1a2темно-серые | A1a1A2A2темно-серые | A1a1A2a2серые | |
| A1a2 | A1A1A2a2темно-серые | A1A1a2a2серые | A1a1A2a2серые | A1a1а2a2светло-серые | |
| a1A2 | A1a1A2A2темно-серые | A1a1A2a2серые | a1a1A2A2серые | a1a1A2a2светло-серые | |
| a1a2 | A1a1A2a2серые | A1a1a2a2светло-серые | a1a1A2a2светло-серые | a1a1a2a2желтые |
Черные чешуйки семян у овса — 1/16, темно-серые чешуйки семян у овса — 4/16, серые чешуйки семян у овса — 6/16, светло-серые чешуйки семян у овса — 4/16, желтые чешуйки семян у овса — 1/16. Расщепление по фенотипу 1:4:6:4:1.
Некумулятивная полимерия имеет место при наследовании формы плодов пастушьей сумки.
| P | A1A1A2A2треугольные | × | a1a1a2a2овальные |
| Типы гамет | A1A2 | a1a2 | |
| F1 | A1a1A2a2треугольные, 100% | ||
| P | A1a1A2a2треугольные | × | A1a1A2a2треугольные |
| Типы гамет | A1A2 A1a2 a1A2 a1a2 | A1A2 A1a2 a1A2 a1a2 |
| A1A2 | A1a2 | a1A2 | a1a2 | ||
| A1A2 | A1A1A2A2треугольные | A1A1A1a2треугольные | A1a1A2A2треугольные | A1a1A2a2треугольные | |
| A1a2 | A1A1A2a2треугольные | A1A1a2a2треугольные | A1a1A2a2треугольные | A1a1а2a2треугольные | |
| a1A2 | A1a1A2A2треугольные | A1a1A2a2треугольные | a1a1A2A2треугольные | a1a1A2a2треугольные | |
| a1a2 | A1a1A2a2треугольные | A1a1a2a2треугольные | a1a1A2a2треугольные | a1a1a2a2овальные |
Треугольная форма плодов у пастушьей сумки — 15/16, овальная форма плодов у пастушьей сумки — 1/16.
Плейотропия — множественное действие генов. Плейотропное действие генов имеет биохимическую природу: один белок-фермент, образующийся под контролем одного гена, определяет не только развитие данного признака, но и воздействует на вторичные реакции биосинтеза других признаков и свойств, вызывая их изменение.
Плейотропное действие генов впервые было обнаружено Г. Менделем, который установил, что у растений с пурпурными цветками всегда имелись красные пятна в пазухах листьев, а семенная кожура была серого или бурого цвета. То есть развитие этих признаков определяется действием одного наследственного фактора (гена).
У человека встречается рецессивная наследственная болезнь — серповидно-клеточная анемия. Первичным дефектом этой болезни является замена одной из аминокислот в молекуле гемоглобина, что приводит к изменению формы эритроцитов. Одновременно с этим возникают нарушения в сердечно-сосудистой, нервной, пищеварительной, выделительной системах. Это приводит к тому, что гомозиготный по этому заболеванию ребенок погибает в детстве. Причиной синдрома Марфана является доминантная мутация гена, контролирующего одновременно рост, длину пальцев, формирование интеллекта и форму хрусталика. Для человека с этим синдромом характерен комплекс следующих признаков — высокий рост, очень длинные гибкие («паучьи») пальцы, повышенный интеллект, близорукость.
Плейотропия широко распространена. Изучение действия генов показало, что плейотропным эффектом, очевидно, обладают многие, если не все, гены.
Таким образом, выражение «ген определяет развитие признака» в значительной степени условно, так как действие гена зависит от других генов — от генотипической среды. На проявление действия генов влияют и условия окружающей внешней среды. Следовательно, генотип является системой взаимодействующих генов.
-
Перейти к лекции №19 «Генетика пола»
-
Перейти к лекции №21 «Изменчивость»
-
Смотреть оглавление (лекции №1-25)
Кодоминирование и множественный аллелизм

Аналогичные расщепления характерны и для кодоминирования с той разницей, что потомки от скрещивания гомозигот разных типов будут характеризоваться наличием фенотипических признаков обоих родителей одновременно. То есть в этом случае оба аллеля являются доминантными: образуют функциональные продукты, которые немного отличаются по некоторым характеристикам. Такая ситуация довольно часто наблюдается для ферментов, когда два аллеля обуславливают синтез двух белков — изоферментов — с практически одинаковой активностью, но при этом они отличаются молекулярной массой (один из белков имеет дополнительный структурный домен и так далее).
Эти примеры требуют важного уточнения. Поскольку в двух гомологичных хромосомах могут находиться только два разных аллеля одного гена, ещё не означает, что в группе особей ген обязательно имеет два аллеля
На самом деле, в большинстве случаев гены существуют в виде нескольких (теоретически неограниченное количество) разных аллелей. Это явление получило название множественный аллелизм.






Одним из примеров множественного аллелизма является система групп крови АВ0. Хорошо известны четыре группы крови — 0, А, В и АВ — определяются тремя аллелями одного гена: ІА, ІВ, і0. Аллели ІА и ІВ являются кодоминантными (отвечают за формирование на поверхности эритроцитов антигенов двух типов — А и В), но при этом доминируют над аллелем і0, который не продуцирует ни одного антигена.
Экспрессия гена
Пенетрантность и экспрессивность. Норма реакции. Экспрессия генов — это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт — РНК или белок. Экспрессия генов может регулироваться на всех стадиях процесса: и во время транскрипции, и во время трансляции, и на стадии посттрансляционных модификаций белков.
Регуляция экспрессии генов позволяет клеткам контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации. Экспрессия генов является субстратом для эволюционных изменений, так как контроль за временем, местом и количественными характеристиками экспрессии одного гена может иметь влияние на функции других генов в целом организме.
Экспрессивность и пенетрантность Под экспрессивностью понимают степень выраженности определенного признака.
Внешняя среда и гены-модификаторы могут изменить экспрессию гена, т. е. выражение признака. Изменчивость проявления мутантного гена у разных особей — довольно частое явление. Например, у потомства дрозофилы — мутантных «безглазых» мух с сильно редуцированным количеством фасеток — содержание их варьирует от почти полного отсутствия до половины нормы. Пенетрантность гена — это доля особей, у которых проявляется ожидаемый фенотип.
При полной пенетрантности (100 %) мутантный ген проявляет свое действие у каждой особи. При неполной пенетрантности (меньше 100 %) ген проявляется фенотипически не у всех особей. Экспрессивность и пенетрантность гена в значительной степени зависят, по-видимому, от влияния генов-модификаторов и условий развития особей.
Норма реакции — способность генотипа формировать в онтогенезе, в зависимости от условий среды, разные фенотипы.
Она характеризует долю участия среды в реализации признака и определяет модификационную изменчивость вида.



